Musa acuminata
Musa acuminata L. A. Colla, Memoria della Reale Accademia delle Scienze di Torino 25 : 384 (1820). [Memoria sul genere Musa e monografia del Medesimo 66 (1820).] and E. E. Cheesman, Kew Bulletin 3 (1): 22 (1948).
| Accepted name | Musa acuminata L. A. Colla, Memoria della Reale Accademia delle Scienze di Torino 25 : 384 (1820). [Memoria sul genere Musa e monografia del Medesimo 66 (1820).] and E. E. Cheesman, Kew Bulletin 3 (1): 22 (1948). |
| Synonyms | Synonyms attributable to subspecies of Musa acuminata: Musa banksii F. J. H. von MuellerMusa microcarpa O. Beccari Musa sumatrana O. Beccari Musa truncata H. N. Ridley Musa zebrina J. E. Planchon Musa zebrina L. B. van Houtte ex J. E. Planchon Synonyms attributable to cultivated seedless forms derived either from Musa acuminata alone or through hybridisation with Musa balbisiana: Musa cavendishii A. B. Lambert ex J. PaxtonMusa cavendishii J. Paxton Musa chinensis R. Sweet Musa chiliocarpa C. A. B. Backer Musa consociata T. Nakai Musa corniculata J. de Loureiro Musa humilis G. (G. G.) S. Perrottet Musa megalocarpa T. Nakai Musa minor T. Nakai Musa mirabilis T. Nakai Musa nana auct. non J. de Loureiro Musa prematura T. Nakai Musa rhinocerotis W. S. Kurz Musa x sapientum L. Musa simiarum var. sylvestris W. S. Kurz Musa simiarum G. E. Rumphius Musa sinensis P. A. Sagot ex J. G. Baker |
| Authorities | The accepted name is from Cheesman 1948b.
The World Checklist of Monocotyledons lists Musa acuminata Colla, Mem. Gen. Musa: 66 (1820) as an accepted name. |
| Section | Eumusa (Musa) 1 |
| Distribution | South East Asia to Australia (very doubtfully Africa). |
| Description | "Plant stooling sparsely (1 - 2 stems, Ridley) or freely (4 - 30 stems, Backer) ; pseudostems more slender than those of most cultivated bananas, usually 3 - 5 metres high (up to 7 m., Backer), rarely exceeding 25 cm. in diameter at base, with varying development of brown-black markings from almost green to almost entirely black, or sometimes reddish brown in lower parts ; leaf-sheaths and petioles commonly more or less glaucous or pruinose, but extremely variable in development of wax. Leaf blades oblong, 2 - 2.5 m. long, 40 - 60 cm. wide, truncate at apex, usually rounded at base, sometimes rounded on one side and acute on the other, varying from green or green tinged with purple to wholly purple on the lower surface, green above, with or without flecks or bars of purplish brown pigmentation, variable also from not or slightly to heavily waxy beneath, midribs green, greenish yellow, or more or less strongly tinged with red below ; petioles 60 - 90 cm. long, in some forms very slender, in others stouter, their margins sometimes almost erect, leaving an open adaxial channel, in other forms strongly incurved over the channel and almost covering it, usually definitely developed, especially below, where the petiole passes into the leaf-sheath, here usually closely appressed to the pseudostem, occasionally slightly bent outward away from it, early becoming scarious in this region, when young often bordered with a red line. Inflorescence subhorizontal or vertically deflexed, its peduncle and rachis usually more or less thickly pubescent with brown hairs, sometimes glabrous ; basal flowers female, the number of female "hands" varying up to about 10, commonly fewer, occasionally more, upper hands male. Female flowers about 16 per bract in two rows ; compound tepal about 2.5 cm. long (to 4.2 cm., Backer) white, yellowish or slightly purple, with white or yellow tip and lobes ; free tepal translucent, about half as long as the compound tepal ; ovary pale green, yellowish green, or purplish, glabrous or with a few fine hairs near the base, or (fide Backer) more or less thickly covered with soft hairs. Male bud in advanced blooming ovoid to turbinate, usually acute, the bracts convolute, imbricate at the extreme tip only, or rather strongly imbricate. Bracts various shades of purple or red, from bright red to dark violet, ovate, usually acute at apex, sometimes yellow at the extreme tip ; outer surface more or less glaucous, faintly ribbed longitudinally, inner surface paler, light red or yellowish, always paling towards the base. Only one bract lifted at a time, bracts revolute on fading and early deciduous, often a little before the flowers they subtend. Male flowers about 20 per bract, in two rows ; compound tepal 3.5 - 4.5 cm. long, about 1.2 cm. wide, white, cream, yellowish or pale orange, sometimes purplish, upper part of tepal including the teeth, yellow, varying from pale lemon yellow to bright orange, the 2 outer teeth varying from 2 to 7 mm. long, with a dorsal filiform appendage 1 - 2 mm. long, the centre lobe usually slightly shorter and broader ; free tepal about half as long as the compound tepal, translucent, boat-shaped, variable at apex but usually transversely corrugated just below the apicula ; stamens at first as long as the perianth or slightly shorter, later exserted, their anthers usually pink before dehiscence. Fruit bunch asymmetrical if borne subhorizontally, compact if vertically, the fruits exhibiting marked geotropic curvature in either case. Individual fruit 8 - 13 cm. long, 1.5 - 3 cm. in diameter, subcylindrical, the angles of the young fruit almost disappearing at ripeness in most forms, rather abruptly narrowed at base into a pedicel of about 1 cm. (sometimes pubescent), and at apex into a prominent acumen 0.6 - 1.5 cm. long ; pericarp about 2 mm. thick, bright yellow at full ripeness, pulp whitish or cream to yellow. Seeds (when present) dull black, smooth (ex Backer) or more commonly minutely tuberculate, irregularly angulate-depressed, 6 - 7 mm. across and about 3 mm. high". (Cheesman 1948b. References to Ridley and Backer in the above description are cited below). |
| References | APNI, Argent 1984, Backer 1924 : 135-136, Backer & Bakhuizen 1968 : 38, Cheesman 1948b, Flora Guandong, Griffiths 1994, GRIN, Hotta 1989, Huxley 1992, IPGRI, Jones 2000 : 10, Mobot VAST database, Ridley 1893, Ridley 1902, Ridley 1924, Rossel 1998, Shepherd 1990, Shepherd 1999, Simmonds 1956 : 469, Uphof 1968, WCM. |
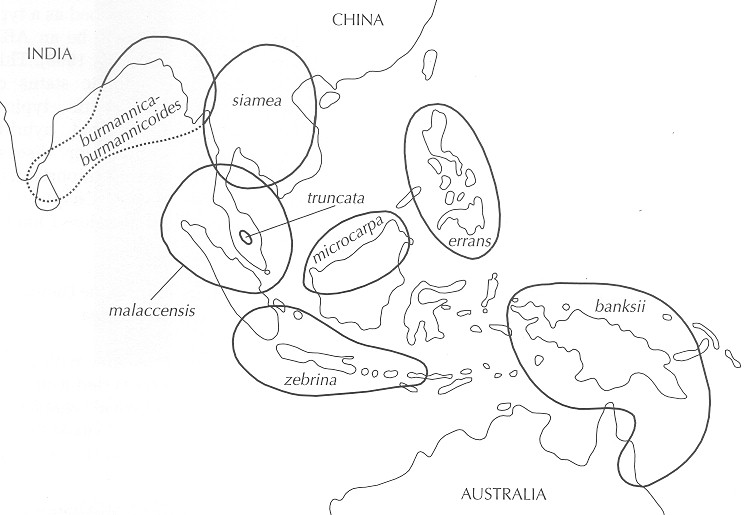
| Comments | Musa acuminata Colla is a highly polymorphic species and its nomenclature is currently in a right old mess. A revision of Musa acuminata is required. If a species is polymorphic it is common for taxonomists to designate intraspecific taxa of which the ICBN allows five possibilities that listed in order of rank are subspecies, varietas, subvarietas, forma and subforma. In the case of Musa acuminata taxonomists have used the intraspecific taxa subspecies, varietas and forma. "Subspecies is used for geographical races, variety is used for local populations which are morphologically and possibly ecologically distinct and the form is used for atypical individuals within a population". The choice of a subspecific designation as opposed to a varietal designation is somewhat erratic in practice. However, there is a hierarchy operating here so that a varietas is a lesser thing than a subspecies. In these circumstances the designation of a variety without a subspecies requires explanation. That explanation is for the author of the name, not me. It is notable that Norman Simmonds states his reasons for choosing subspecific rank for the various forms of M. acuminata in his 1956 paper. Such reasoning is usually not given. For example, Häkkinen and Wang Hong (2007) give no explanation for why they choose varietal status for Musa acuminata var. chinensis as opposed to subspecific status which they maintain for another Chinese form of Musa acuminata viz. subsp, burmannica. Twenty three (23) intraspecific taxa of Musa acuminata are listed on subsequent pages, subspecies and varietas and forma but there is no formal hierarchy as might reasonably be assumed. It is likely that not all taxa are valid. Authorities for some of the taxa are currently lacking in this list. The principal subspecies are indicated on this distribution map for Musa acuminata from Jones 2000.
The relationship between certain of the sub-species of Musa acuminata is somewhat confusing especially in relation to the horticultural 'forms' of the subspecies in cultivation. These tend to be given different names by different nurseries or different literature e.g. ‘Bordelon’, 'Sumatrana', 'Zebrina', 'Rubra' and 'Rojo'. The names are used sometimes as if they were species names. This is wrong. The names are used sometimes as if they were cultivar names. This is also probably wrong in a formal sense because most are probably referable to one subspecies (or variety) or another. However, some plants may indeed be distinct selections worthy of cultivar status. Unless and until someone collects all the cultivated 'forms' together in one place and evaluates them side-by-side it is likely not to be possible to determine their true individual status. Musa acuminata is an Australasian species although there is one peculiar population in the forests north of Wete on Pemba Island off the Tanzanian coast of east Africa. Stover & Simmonds 1987 speculated that Musa acuminata may be wild on Pemba Island in which case it would be the only wild Musa species with a foothold in Africa. It is more likely that the plant was introduced to Pemba from Indonesia, see for example Rossel 1998, although Shepherd 1999 treats it as a possibly distinct subspecies. |
last updated 30/04/2008